Presented at the 7th Annual Computational Neuroscience Meeting (CNS*98), Santa Barbara (CA), July 26-30th 1998
Figure-ground segregation of coherent motion in V1: A model based on the role of intra-cortical and extra-cortical feedbacks
Email:
An important use of visual motion is in the segregation of a complex scene into
its perceptually coherent and distinct moving components. The recent
computational theories have expressed the perception of coherent visual motion
as an optimization process constrained by the conflicting demands of
integration and segmentation of local motion [1-2]. They tell us however very few about the nature of the
cortical mechanisms involved in our spatially accurate perception of coherent
motion. Recent anatomical and physiological experiments have however provided
strong evidence that V1 processing is under the influence of contextual
modulations [3] through intra- or extra-cortical
connections [4]. They also suggested its perceptive
correlate in terms of figure-ground segregation as soon as area V1 [5], a view clearly opposite to the hierarchical functional
organization of the visual system [6].
This work investigates the function of the interactions between visual areas V1
and MT in the figure-ground segregation of coherent motion, with the aim of
revealing the nature of the integrative processes performed by area V1 in
segmentation of visual motion.
A neuromorphic model of the retino-cortical motion processing stream is
proposed which incorporates both feedforward and feedback mechanisms. The
feedforward stream follows a gradient scheme for local motion estimation and
its integration for global motion estimation, while the feedback stream applies
local constraints of both smoothness and discontinuity onto the local motion
stage. The functional properties of this model were derived analytically, and
are illustrated by real-time simulations on image sequences representing real
scenes and psychophysical stimuli.
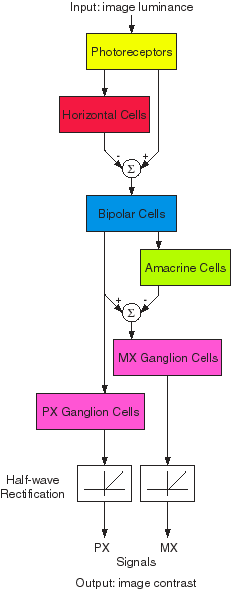
Structure of the model.
Each neural layer of the model (6 in retina, 3 x n in V1, n in MT where n is
the number of considered orientations) is modeled by a resistive network of
leaky integrators. Spatial and temporal characteristics of the neurons are
reflected by the coupling between neighbor units, and their membrane
properties, respectively. Synaptic interactions are considered as linear for
the excitatory or inhibitory ones and as multiplicative for the modulatory
ones.
Plausibility of the model.
We propose that this model correlates with the following neural organization of
the visual pathway:
|
i) Retinal spatiotemporal processing provides parvo- and magno-cellular inputs with different band-pass characteristics to area V1.
ii) Oriented band-pass filtering of the luminance parvo-cellular signal provides V1 orientation-selective simple cells responses.
iii) These cells interact nonlinearly with the magno-cellular inputs to produce V1 direction-selective simple cells (SDS) responses.
iv) These V1 SDS cells drive the V1 direction-selective complex cells (CDS) which are laterally and excitatory connected.
v) The spatial convergence and temporal integration of V1 CDS signals provide the responses of MT direction-selective cells with large receptive fields.
vi) The reciprocal retinotopic connections from MT modulate the gain of the feedback loop formed by the excitatory intra-cortical connectivity between V1 CDS cells. |
Retino-cortical model of the motion processing stream |
Simulations.
The striking capabilities of the model are illustrated on the processing of
kinetic contours defined by coherent random-dot motion: a spatial shape
composed of coherent moving random-dots embedded in a background composed of
non coherent moving random-dots is successfully segmented by the V1 CDS cells
in presence of the MT feedback, while the deactivation of this feedback leads
irremediably to the disappearance of this shape. The feedback onto V1 also
increases significantly the signal-to-noise ratio at the level of area MT, and
speeds up its activation in presence of coherent motion.
Retinal Stage |
 |

The region surrounded by the dashes only delineates the spatial shape formed by 50% of coherently moving random-dots.
|
Cortical Stage |
 |



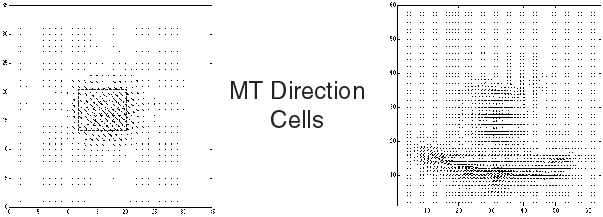
|
Sorry for the low quality of figures with arrows representing estimated motion in the different layers, I will improve these as soon as possible, as well as to add some QuickTime movies showing temporal simulations of the model.
|
This particular scheme has three important consequences on the representation
of visual motion:
1) Area MT imposes a local smoothness constraint onto the representation of the
optical flow by V1 CDS cells, and the aperture problem is solved through the
selection of the optimal receptive field which integrates the local motion in a
way consistent with the global estimation;
2) The overall effect of this space-variant alteration of the receptive fields
is an effective enhancement of V1 CDS responses at locations where the local
motion is compatible with the global motion, and a dramatic decrease where it
is not. This spatially nonlinear mechanism then preserves the discontinuities
present in the "true" motion across space, and thus promotes a
shape-from-motion segmentation as early as area V1;
3) This figure-ground segregation of coherent motion is the result of an
optimization process which converges in presence of stationary motion, and
which is supported by the dynamics induced through the reciprocal connections
between areas V1 and MT.
The explanation of this result rests on the specific nature of the feedback
made by area MT onto area V1 in the proposed model. Indeed, MT feedback onto V1
does not just transmit a re-entrant signal but rather an active signal which
modulates the V1 processing according to a confidence measurement of motion
coherence. Responses of MT cells increase in presence of coherent motion, and
then favour the spatial and temporal integration of local motion through the V1
CDS intra-cortical network. Conversely in absence of coherent motion the weaker
responses of MT cells limit such integration to the direct input from V1 SDS
cells which carry the local estimation. The enhancement or depression of V1 CDS
responses according to the strength of MT responses reflects these changes in
the spatiotemporal properties of V1 CDS receptive fields (gain, space and time
constants).
We proposed a neuromorphic model of the motion pathway based on these cortical
mechanisms. The feedforward stream alone provides a spatially "fine"
representation of local motion in area V1 and a spatially "coarse"
representation of global motion in area MT. It is however unable to deal with
the aperture problem, and consequently cannot support any spatial segmentation
based on coherent motion. Reciprocally the feedback from MT onto V1 induces a
dynamic competition between the local and global representations of motion
information, which promotes the emergence of an intermediate representation of
the visual flow compatible with both local and global representations. On
convergence of this dynamics, responses of V1 CDS cells provide a fine
representation of "true" motion, solving simultaneously the aperture problem
and the figure-ground segregation of coherent motion.
The perception of coherent visual motion can be expressed as an optimization
problem constrained by the conflicting and simultaneous demands of integration
and segmentation of local motion, and requires a nonlinear regularization
approach [7]. By implementing such a computational scheme,
the proposed model accounts for the figure-ground segregation of coherent
motion as soon as area V1 through the modulation of its intra-cortical
connections by MT feedback. The model is also compatible with the recent
anatomical and physiological evidence of contextual modulation inside area V1
[3-5], and:
1) It demonstrates that area V1 is able to subtend the figure-ground
segregation of coherent motion with a high resolution spatial information;
2) It suggests, by the simplicity of its neural structure, that the excitatory
intra-cortical connectivity and its modulation by a higher cortical signal is a
critical feature of the cortical circuitries involved in visual segmentation;
3) Although initially based on a theoretical background and developed in the
context of neuromorphic engineering, the model also provides some
experimentally testable hypothesis about the nature of the integrative
processes performed by area V1 as suggested by the recent theories [8-9].
1. Poggio, Torre & Koch. Computational vision and
regularization theory. Nature 317, 314-319 (1985).
2. Yuille & Grzywacz. A computational theory for the
perception of coherent visual motion. Nature 333, 71-74 (1988).
3. Zipser, Lamme & Schiller. Contextual modulation in
primary visual cortex. J. Neurosci. 16:7376-7389 (1996).
4. Hupé, James, Girard & Bullier. Feedback connections
from V2 modulate intrinsic connectivity within V1. Soc. Neurosci. Abs.
406.15 (1997).
5. Lamme, van Dijk & Spekreijse. Contour from motion
processing occurs in primary visual cortex. Nature 363, 541-543 (1993).
6. Bullier & Nowak. Parallel versus serial processing:
new vistas on the distributed organization of the visual system. Current
Opinion in Neurobiology 5, 497-503 (1995).
7. Schnorr & Sprengel. A nonlinear regularization
approach to early vision. Biological Cybernetics 72, 141-149 (1994).
8. Mumford. On the computational architecture of the
neocortex: II The role of cortico-cortical loops. Biological Cybernetics
66, 241-251 (1992).
9. Lee, Mumford, Romero & Lamme. The role of the primary
visual cortex in higher level vision. Vision Research 38, 2429-2454
(1998).
© 1998 KyberVision Consulting, R&D
E-mail:
